|
О
существовании биологических ритмов
людям известно с древних времен.
Уже в
«Ветхом Завете» даны точные указания о правильном образе жизни, питании, чередовании фаз активности и отдыха. Об
этом же писали многие выдающиеся ученые древности: Гиппократ, Авиценна и
другие.
Первые
серьезные научные исследования в этой области начали проводиться в начале ХХ века, в том числе И.П.
Павловым, В.В. Вернадским, А.Л. Чижевским и другими учеными.
Большое
число работ было посвящено исследованиям различных параметров биологических
ритмов, их связи с внешней средой, методам лечения и профилактики различных
заболеваний.
В
качестве примеров можно привести исследования воздействия света на циркадные
колебания организма («Правило Ашоффа»), исследования суточных ритмов
биологических процессов и их адаптивного значения в онто-и филогенезе
позвоночных (Губин, Герловин 1980), изучение влияния биоритмов на
работоспособность (Агаджанян Н.А и др. 1978) и т.п.
Особые
усилия ученых были направлены на поиски так называемых «биологических часов» - циркадного центра.
В результате в 1972 году двум группам
американских исследователей (группе Роберта Мура - Виктора Эйхлера и группе
Фредерика Стефана - Ирвина Цукера) удалось доказать, что центром управления
биологическими часами организма является супрахиазматическое ядро.
Кроме
того, вице-президентом международного общества хронобиологов, профессором
Миннесотского университета США Францем Хальбергом была разработана классификация биологических ритмов.
В
соответствии с этой классификацией биоритмы делились следующим образом:
Ультрадианные
ритмы с периодом меньше 20 часов, циркадианные - с периодом 24±4 часов и
инфрадианные - с периодом больше 28 часов.
Среди
последних были выделены: циркасемисептанные ритмы с периодом примерно 3±0,5
сут; циркасептанные ритмы с периодом 7±3 сут., циркадисептанные - с периодом
14±3 сут., циркавигинтанные - с периодом 21±3 сут., циркатригинтанные - с
периодом 30±5 сут., цирканнуальные - с периодом 1 год ± 2 месяца.
К
концу 20-го века факт ритмичности биологических процессов живых организмов стал считаться одним из
фундаментальных свойств живой материи и сущностью организации жизни (Комаров,
Рапопорт, 2000).
Однако,
несмотря на значительные достижения в исследованиях биологических
ритмов, до сих пор не установлена их
причина.
Целью
настоящей работы является определение причины периодичности физиологических
процессов – биологических ритмов и их роли в процессе жизнедеятельности организма.
Для
определения причины периодичности физиологических процессов нами были
рассмотрены термодинамические особенности функционирования живых организмов.
Известно,
что одним их основных условий жизнедеятельности живых организмов является
необходимость их энергетических
затрат. Причем получение свободной энергии из продуктов питания и ее потребление живыми организмами
осуществляется с помощью метаболизма – циклов обмена веществ, непрерывно
происходящих в клетках этих организмов (Северин, 2003).
Необходимость энергетических затрат
живыми организмами впервые были объяснены Эрвином Бауэром в начале 20-го
века, сформулировавшем «Всеобщий
закон биологии» в следующей редакции (Бауэр, 2002):
|
«Все и только живые
системы никогда не бывают в равновесии и исполняют за счет своей свободной
энергии постоянную работу против равновесия, требуемого законами физики и
химии при существующих внешних условиях».
|
Э. Бауэром
также был сформулирован «Принцип
устойчивого неравновесия живых систем»:
|
«Для живых систем
характерно именно то, что они за счет своей свободной энергии производят
работу против ожидаемого равновесия».
|
В этой формулировке под
«ожидаемым равновесием» Э.Бауэром понималось равновесие, к которому стремится
неживая природа в условиях замкнутых систем в соответствии с законами
равновесной термодинамики (второе начало термодинамики).
Позже идея Э. Бауэра была
подтверждена И. Пригожиным в его фундаментальных работах по неравновесной
термодинамике (Пригожин, 1986). Им было установлено, что вся окружающая нас
материя (в том числе и живые организмы) всегда находится в неравновесных
термодинамических состояниях в условиях открытых систем.
Однако Э. Бауэр,
теоретически предположив наличие структур, обеспечивающих указанное
неравновесие, не раскрыл сущности их функционирования.
Он лишь
утверждал, что живые системы непрерывно производят работу против ожидаемого
равновесия, то есть постоянно и в одном направлении.
В связи с
этим нами были проведены
исследования, направленные на определение механизмов,
обеспечивающих состояние устойчивого
неравновесия живых систем, а так же закономерности их работы, и в частности
анализ процессов обмена веществ, происходящих в клетках (метаболизм).
Выполненные работы показали
следующее.
Как
известно, процессы метаболизма представляют собой комплексы разнообразных
циклов биохимических реакций расщепления и синтеза веществ, происходящими в
клетках ( Северин 2000 ).
При этом в клетках
происходят непрерывные периодические колебания концентраций веществ,
участвующих в этих реакциях, например
биохимические реакции с участием кальция (рис. 1), биохимические реакции
гликолиза и др.
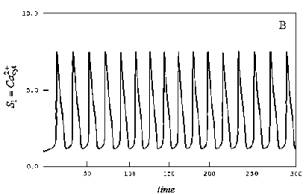
Рис.1.
График внутриклеточных колебаний кальция. (Dupont, Goldbeter, 1983).
Также
известно, что основным носителем энергии в живых организмах является аденозинтрифосфат (АТФ), причем циклы
синтеза-расщепления АТФ лежат в основе функционирования отдельных белков,
белковых комплексов, клеточных структур, органелл и клеток в целом (Горбачев,
2000; Северин, 2003).
Поскольку
практически для любого проявления жизнедеятельности клеток необходим АТФ,
основная метаболическая активность живых клеток направлена на его синтез.
Синтез АТФ
происходит путем различных сложных последовательностей биохимических реакций,
в которых используется потенциальная химическая энергия, заключенная в
молекулах углеводов и жиров (Северин, 2003).
Расщепление
АТФ может происходить постадийно путем гидролиза с выделением свободной
энергии в виде следующих реакций (Горбачев, 2000):
|
АТФ + H2O → АДФ + Фн+
H+
+ ∆G = -30 кДж/моль
АДФ + H2O → АМФ + Фн+
H+ +
∆G = -30
кДж/моль
АМФ + H2O → аденозин
+ Фн + H
+ ∆G = -13
кДж/моль
|
Здесь:
АТФ –
аденозинтрифосфат
АДФ – аденозиндифосфат
АМФ - аденозинмонофосфат
H+ -
положительный ион водорода,
Фн - неорганический фосфат
∆G - изменение
свободной энергии
Однако
обычно клетки извлекают энергию из АТФ во время клеточного
дыхания, при котором происходит только первая стадия расщепления АТФ: от его
молекулы отщепляется только одна фосфатная группа.
В процессе
синтеза-расщепления АТФ его концентрация в клетках непрерывно меняется. В
упрощенном виде пример закономерности изменения концентрации АТФ в клетке в
процессе непрерывных циклов синтеза-расщепления АТФ может быть представлен в
виде графика, изображенного на рис.2.
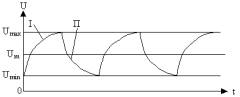
Рис.2. Типовой
график синтеза-расщепления АТФ.
U
- величина концентрации АТФ в
клетке;
I – процесс синтеза; П – процесс
расщепления.
Как видно из графика на рис.2., процессы синтеза и
расщепления АТФ происходят по законам, близким к экспоненциальным. При этом
верхние и нижние границы концентрации биохимических веществ и скорость
протекания биохимических реакций определяются соответствующими положительной
и отрицательной обратными связями, имеющимися в живых системах. Их роль
выполняют соответствующие ферменты -
биохимические катализаторы и ингибиторы, ускоряющие или замедляющие эти
реакции (Северин, 2003).
Пример
схемы работы механизма регуляции ферментативных реакций представлен на рис.
3.
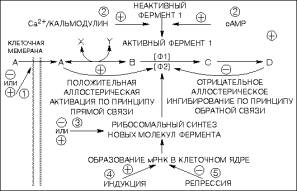
Рис. 3. Механизмы
регуляции ферментативных реакций
|
Цифры, заключенные в кружки,
указывают вероятные участки действия гормонов. 1 - изменение проницаемости
мембраны; 2 - переход фермента из неактивной формы в активную; 3 -
изменение скорости трансляции мРНК на рибосомальном уровне; 4 - индукция
образования новой мРНК; 5 - репрессия образования мРНК.
|
Аналогично
происходят и групповые когерентные биохимические реакции на уровне органов,
систем и организмов в целом, вызывая
у последних соответствующие функциональные сдвиги (Ризниченко, Рубин 1993).
На рис.4.
представлен пример идеализированных типовых колебаний функциональных сдвигов организма человека
по результатам исследований проф. В.М.Ретнева (Ретнев, 1984). В процессе этих
исследований круглосуточно, каждые четыре часа в течение недели
контролировались различные физиологические параметры водителей общественного
транспорта: частота сердечных сокращений, температура тела, систолическое и
диастолическое артериальное давление, реакции на свет и на звук. Как показали результаты этих исследований,
закономерность изменений функциональных сдвигов этих физиологических
параметров носила идентичный характер, с выраженным периодом колебаний в одни
сутки и одну неделю.
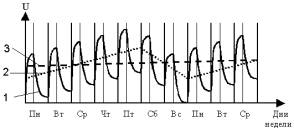
Рис.4. Пример типовых колебаний функциональных
сдвигов организма
водителей общественного
транспорта.
1 –
суточные колебания функционального сдвига;
2 –
недельное среднедействующее колебание функционального сдвига;
3 –
среднедействующее значение функционального
сдвига.
Анализируя эти процессы с точки зрения биологии, можно убедиться,
что колебания функциональных сдвигов
действительно представляют собой результат когерентных непрерывных
биохимических реакций последовательностей метаболических циклов, в основном
содержащих чередующиеся комплексные
процессы, направленные на синтез и
расщепление АТФ. В результате этих реакций все параметры внутренней среды
живых организмов находятся в состоянии непрерывных колебаний относительно
соответствующих средних значений.
Однако с точки зрения физики эти процессы представляют собой
когерентные непрерывные
термодинамические колебания, содержащие фазы соответственно затрат и
выделения энергии. Размах и частота этих колебаний регулируются существущими
в биологических системах обратными связями, упомянутыми выше, а количество
выделяемой энергии всегда больше затрачиваемой.
По принципу действия эти процессы могут быть представлены в виде
работы биохимического двухтактного импульсного генератора энергии, содержащего
непрерывно чередующиеся такты ее выделения и потребления.
Для иллюстрации этих процессов на рис. 5 представлены
графики:
а) чередования синтеза и
расщепления АТФ;
б) чередования затрат и
выделения энергии.
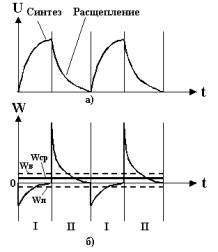
Рис.5. а) График чередования
синтеза и расщепления веществ;
Рис.5. б) График чередования затрат
и выделения энергии;
Wп – среднедействующее значение
потребляемой энергии при синтезе АТФ;
Wв – среднедействующее значение выделяемой энергии при
расщеплении АТФ;
Wср – среднедействующее
результирующее значение выделяемой энергии
Как видно из графиков рис.5,а) и б), циклы биохимических реакций
синтеза и расщепления АТФ сопровождаются соответствующими циклами
термодинамических колебательных процессов потребления энергии Wп (фаза I) и выделения энергии Wв (фаза II).
В результате совокупного действия указанных циклов колебательных
термодинамических процессов выделяется энергия:
Wср=Wв – Wп > 0.
Эти непрерывные термодинамические колебательные процессы,
посредством которых выделяется энергия, и являются механизмом обеспечения
устойчивого термодинамического неравновесия.
Таким образом, в дополнение к первому всеобщему закону биологии,
можно сформулировать второй, изложенный в следующей редакции:
|
Устойчивое
термодинамическое неравновесие в живых системах обеспечивается непрерывными
чередующимися фазами выделения и потребления энергии, причем количество
выделяемой энергии всегда больше потребляемой.
|
Поскольку каждый живой организм по своему уникален, он всегда имеет
только ему принадлежащий набор
параметров амплитуд и частот периодических колебаний в его клетках, органах,
системах и организме в целом.
Причем параметры этих колебаний постоянно меняются и зависят от самых
разных причин: состава и качества пищи, условий жизни, состояния организма,
возраста, окружающей среды и т.д. Это
же в полной мере относится и к организму человека.
Однако окружающая среда, в которой живет человек (как и большое число
других живых организмов), принудительно заставляет его организм жить с
основным периодом колебаний функуиональных сдвигов в одни сутки. Это связано
как с периодичностью дня и ночи, так и с социальными и другими факторами.
В связи с этим организм человека компенсирует разброс между
собственными ритмами и принудительным ритмом продолжительностью сна, образом
жизни, режимом дня и другими средствами.
Полученные результаты исследований были сопоставлены с материалами
различных других работ в области хронобиологии и хрономедицины ( Агаджанян
и др. 1978; Ашофф 1984; Бреус и др. 2002; Губин, Герловин 1980; Гласс, Мэки
1991; Ризниченко, Рубин 1993; Чернух, Гехт 1982 и др.)
Приведенные
в этих работах практические результаты в подавляющем большинстве случаев
совпадают с результатами настоящего исследования.
Например, из приведенного выше графика на рис.4.
видно, что основные периоды колебаний функциональных сдвигов организма
водителей общественного транспорта
составляют как 24 часа, так и
7 суток. Это объясняется тем, что труд водителей является достаточно тяжелым,
в связи с чем после трудового дня за время отдыха до начала следующего дня
функциональные сдвиги, вызванные работой, у них полностью не
восстанавливались и наблюдалось накопление остаточных функциональных сдвигов
в течение рабочей недели. Полное же восстановление функциональных сдвигов
происходило лишь за выходные дни (Бутковская и др. 1998; Доброборский,
Кадыскина 1997, 2000).
Определенную периодичность изменений функциональных
сдвигов можно обнаружить и за годовой период, связанный с временами года и
режимом питания.
Таким
образом, причиной периодичности физиологических процессов (биологических
ритмов) в живых организмах является метаболизм, при котором происходят
когерентные колебания выделения и затрат энергии, за счет разности которых
обеспечивается их устойчивое термодинамическое неравновесие.
На
основании полученных данных можно
сделать следующие выводы:
1. Принципом существования всех живых организмов является
выделение свободной энергии путем непрерывно чередующихся фаз ее выделения и
потребления, при которых величина
выделяемой энергии всегда больше потребляемой. Именно этим путем
обеспечивается их неравновесное термодинамическое состояние.
2. Ни один процесс в живых организмах не может протекать
постоянно, а должен непрерывно чередоваться с обратными: труд с отдыхом, вдох
с выдохом, мышечные сокращения с
расслаблениями, бодрствование со сном
и т.д.
3. Состояние любого живого организма никогда не бывает
статическим, а все его физиологические параметры: пульс, артериальное
давление, температура, состав крови, физиологические реакции, функции нервной
деятельности и др. всегда находятся в состоянии непрерывных колебаний
относительно некоторых средних значений.
СПИСОК ЛИТЕРАТУРЫ
Агаджанян
Н.А., Горшков М.М., Котельник Л.А., Шевченко Ю.В. Ваша работоспособность
сегодня. М. Советская Россия, 1978. – 88 с.
Бауэр
Э. Теоретическая биология. Росток. 2002.
Биологические ритмы / Под ред.
Ю. Ашоффа: В 2 т. - М.: Мир, 1984
Бреус Т.К., Чибисов С.М. ,
Баевский Р.М., Шебзухов К.В. Хроноструктура биоритмов сердца и внешней среды.- М.:
Издательство Российского университета дружбы народов, 2002. — 232 с.
Гласс Л., Мэки М. От часов к хаосу. Ритмы жизни.
- М.: Мир, 1991.
Горбачев
В.В. Концепции современного
естествознания. МГУП, М., 2000.
Губин
Г.Д., Герловин Е.Ш. Суточные ритмы биологических процессов и их
адаптивное значение в онто-и филогенезе позвоночных / АН СССР. Сиб. отд-ние;
Акад. мед. наук СССР. Сиб. филиал. – Новосибирск: Наука, 1980.
Бутковская З.М., Доброборский Б.С., Кадыскина Е.Н. К
вопросу применения нового эргономического показателя Эргоемкость при
гигиенической оценке и сертификации ручного инструмента. Медицина труда и
промышленая экология. №11. 1998. М. Стр 15 – 17.
Доброборский Б.С., Кадыскина Е.Н. К вопросу измерения биологических затрат
человека при
эргономической оценке
промышленного оборудования и промышленных товаров народного потребления.
Тезисы докладов научной конференции «Проблемы санитарно-эпидемиологического
благополучия населения Северо-Западного и других
регионов Российской федерации».
С-Пб., 1997. С. 53-54.
Доброборский
Б.С., Кадыскина Е.Н. Способ количественной оценки воздействия на организм
человека различных нагрузок. Патент Ru № 2159576. Бюллетень Открытия, изобретения, промышленные
образцы, торговые марки. 27.11.2000. №33, С. 127.
Комаров
Ф.И., Рапопорт С.И. Хронобиология и хрономедицина. «Триада-Х». М. 2000.
Пригожин И., Стенгерс И. Порядок из хаоса: Новый диалог
человека с природой. «Наука» М. 1986.
Ретнев В.М. Гигиена труда водителей городского общественного транспорта. «Наука», Л.
1984. 132 с.
Ризниченко Г.Ю., Рубин А.Б. Математические модели
биологических продукционных процессов. М., Изд. МГУ, 1993, 301 с.
Северин Е.С. Биохимия.
«ГЭОТАР-МЕД», М. 2003
Чернух А.М., Гехт Б.М. Биологические
ритмы и колебательные процессы как инструмент анализа фундаментальных проблем
теоретической и практической медицины.- В кн.: Хронобиология и хрономедицина.
Тез. докл. 2-го симпозиума СССР-ГДР, Тюмень, Тюменск. мед. ин-т, 1982.- С.
6-7
|
