
|
<<<
Interlibrary.narod.ru |
|
|
А.М. Королев, С.М. Василенко
ВСЕСОЮЗНАЯ ОРДЕНА ЛЕНИНА И ОРДЕНА ТРУДОВОГО КРАСНОГО ЗНАМЕНИ АКАДЕМИЯ
СЕЛЬСКОХОЗЯЙСТВЕННЫХ НАУК
имени В.И. ЛЕНИНА.ОРДЕНА ТРУДОВОГО
КРАСНОГО ЗНАМЕНИ
АГРОФИЗИЧЕСКИЙ НАУЧНО - ИССЛЕДОВАТЕЛЬСКИЙ ИНСТИТУТ
НАУЧНО – ТЕХНИЧЕСКИЙ БЮЛЛЕТЕНЬ ПО АГРОНОМИЧЕСКОЙ ФИЗИКЕ № 79
ЛЕНИНГРАД 1990
|
УДК
518.1+541.69+519.25 |
МЕТОДЫ МНОГОФАКТОРНОГО АНАЛИЗА В ПРОЦЕДУРЕ
ИДЕНТИФИКАЦИИ БИОХИМИЧЕСКИХ МЕХАНИЗМОВ
С ЦЕЛЬЮ КЛАССИФИКАЦИИ БИОЛОГИЧЕСКИХ ОБЪЕКТОВ
|
Представление о супероптимальных,
или стрессирующих, воздействиях как об инициаторах глубоких биохимических (в
том числе надмолекулярных) перестроек, которые в той или иной мере
затрагивают все клеточные структуры, делает "дозирующий" стресс
привлекательным методом идентификации биологических объектов. Различная инерционность фенотипического и собственно
генотипического откликов позволяет использовать понятие о накопленном
(интегральном) сигнале-” в котором бы
отражались и быстрая
конструктивная перестройка
структур и (или) метаболизма /1,2/ и глубокая адаптация, обусловленная
синтезом белков de - novo. /3,4/. Это сразу позволяет говорить о портрете
явления на базе как минимум пяти численных маркеров: амплитуда и расстояние
между не медленным к отсроченным возмущениями опорного сигнала, гистерезис, максимальные
отклонения (+1; -1). Если дополнительно рассматривать скорость фронтов
импульса (d h / d t) и их изменения (d h 2/ d2 t), идентификационная информативность матрицы резко
возрастает /5,б/. Однако подход, в основу которого
положен анализ одно-, а том более многопараметрического отклика после
однократного воздействия, ограничен по своему внутреннему условию, так как
предполагает последовательную процедуру анализа уже свершившегося явления, начальные
условия которого по существу случайны и неподконтрольны исследователю. Он не
может выйти за пределы естественных системных отношений внутри объекта
тестирования и между ним и окружающей средой. Это даже без учета того, что
многопараметричность предполагает стандартизацию двух и более измерительных
процедур и неизбежную в этой случае машинную (да ЭВМ) обработку совокупности
сигналов. Усложнение аналитической процедуры не должно вступать в
противоречие с требованиями представительности выборки, особенно если речь
идет о микрообъектах с единственной целевой функцией(биосинтетики, например),
а также об объектах со сложным жизненным циклом. Последовательный анализ
большого количества образцов, сосредоточенный на получении максимально
возможного спектра сигналов-откликов, сразу наталкивается на фактор времени.
Причем если лабораторное время всего массива измерений еще можно как-то контролировать,
то собственное биологическое время (пластохрон) анализируемых объектов
остановить невозможно, а учесть затруднительно. Когда говорят об анализе
сигналов, предполагается, что речь идет об изучении последовательного
однократного возмущения. В этом случае смысл многопараметрического съема
после стрессовой информации по всему доступному спектру признаков - эта попытка
аппаратными средствами преодолеть "не специфичность" отклика. Однако,
как нам кажется, есть эффективный процедурный путь преодоления указанных
ограничений. В самом деле, пространству состояний биологической системы -
гомеорезу (по Удингтону)- соответствует пространство среды обитания (принцип
адекватности), тогда любой физиологический стресс для совершенно .разных
биологических объектов на самом деле будет принадлежать системе одних и тех
же элиминирующих факторов. В результате мы имеем картину, когда согласно
принципу наименьшего взаимодействия Гельфанда - Цетлинс” /7/ биологическая
система, минимизируя свое взаимодействие со средой, дает стереотипный отклик,
который является "слепком" среды обитания, а не отражением
глубинной генетической специфичности данной биологической системы. В случае
же запредельных возмущений, когда регистрируется динамика необратимых
разрушений биологической систему мы тем более будем иметь дело
исключительно с фенотипом, а высказанные выше сомнения лишь обостряется. Получается,
что адаптивные реакции вовсе не лучший маркер для идентификации биологического
объекта? Все не так безнадежно. Мы лишь столкнулись с ограниченностью
классического подхода, который характеризуется установлением зависимости не
от множества, но лишь от какого-либо
экстремального фактора, т.е. с точки зрения аналитической процедуры является
однофакторным (ограниченным). Выше уже было сказано, что расширение числа
стрессоров наталкивается на проблему их количественной стандартизации, но
главное- не преодолевает факторного пространства среды обитания биологической
системы. Другое дело, если на период обработки стрессором
"расширять" либо "сужать" факторное пространство среды
обитания. Мы как бы вторгаемся в совокупность факторов отбора (элиминации), и
в этом случае уже на стресоор как таковой, а фактор, изменяющий пространство
отклика, является потенциальным маркером биологической системы. При таком
подходе вполне оправдан переход от одно- к многофакторному зондированию
биологической системы, но. на фоне единственного стрессора. В качестве
примера показаны результаты факторных экспериментов с актиномицином
продуцентом Str.1етогia (штамм 78 противогрибкового антибиотика леворин).
Варьирование факторов производилось согласно ортогональной матрице 4 х 4 /8/.
Выходной параметр "оптимизации активность на сливе. Стрессор - термоэкспозиция при 45 °С в течение 2 ч. Факторное
пространство среды обитания изменялось при помощи энергетического субстрата,
в качестве которого использовали сукцинат натрия. Варьируемые
компоненты подбирались по признаку химического подобия комплексному
органоминеральному источнику питания - кукурузному экстракту. В таблице
представлены расчетные матрицы эффектов. Результаты факторных экспериментов с актиномицином без тепловой обработки (а) и на фоне теплового шока 45 °С (б)
Из
приведенного примера видно, что введение сукцината в физиологических концентрациях
в состав комбинационных вариантов сред заметно усиливает деформацию матрицы
после однотипной обработки всех вариантов культуральной жидкости
супероптимальной температурой. Изменение факторного пространства среды
(присутствие сукцината) приводит к индивидуализации оптика через актуализацию
"числового образа-портрета" явления (см.таблицу, 1-й уровень).
Возможности повышения информативности математического планирования для микробиологических
сред уже обсуждались в работе /9/ и развитие этой темы в данной статье вряд
ли целесообразно. В
приведенных примерах для классификации объекта использовались только входные
параметры, но не описывался объект как таковой. В следующем примере
предложено многопараметрическое описание классифицируемого объекта,
проведенное на фоне факторного пространства по множеству параметров
(дескрипторов). В результате каждому объекту классификации сопосавляется
вектор X, а все объекты могут быть представлены в виде матрицы: Далее
процедура распознавания объектов складывается из масштабирования, нормализации, преобразования
кластеризации /10/, отбора признаков /II/, многомерного скейлинга, нелинейного
отображения /12/ и кластеризации /13/. Реакция
организмов на стрессовые воздействия существенным образом зависит от состояния
клеточных мембран как первичных акцепторов внешнего сигнала /1,2,4/. Поэтому
привлекательно перейти от оценки метаболических последствий стресса к прямому
диагностирования состояния мембранных комплексов. Во многих случаях такая
постановка
задачи
означает поиск способов контроля изменении трансмембранного потенциала
хлоропластов, митохондрий (прокариот или эукариот) /14/ либо сценку изменений активности цитохромоксидазного и
дегидрогеназного комплекса, т.е. ферментных цепей, в которых происходит
биотический перенос электронов. Также мы
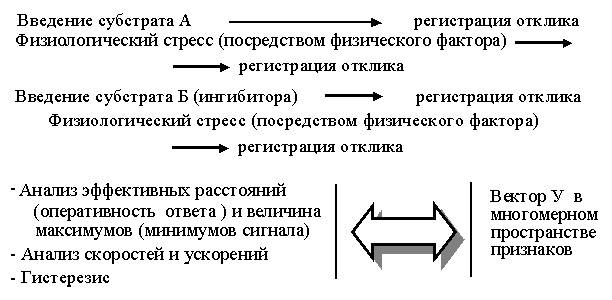
считаем целесообразным переход от матричного планирования (т.е. набора
исходно независимых процедур) к последовательному изменению факторного
пространства среды по типу:
Приборное
обеспечение последовательной процедуры, построенной в принципе на необратимом
разрушении объекта, формируем в зависимости от видоспецифичности биологических
объектов. Так, для микроорганизмов аэробов и некоторых факультативных видов
опорный сигнал легко получить, регистрируя способность культур к
электрогенезу при их инкубации в биохимических топливных элементах;
для семян (зародышей) растений это может быть последовательная хроматография
газов при инкубации последних в замкнутом пространстве; для листьев,
водорослей, хлоропластов - замедленная флюоресценция и т.д. В
заключение еще раз о главной мысли работы. Авторы считают, что искусственно изменяя
факторное пространство среды за счет введения экзотических доноров и
акцепторов электронов на момент воздействия агрессоров на биологическую
систему, можно преодолеть "не специфичность" физиологического
(биохимического) отклика и использовать форму регистрируемого сигнала (по
любому доступному, желательно энергетической природы, параметру) для
высокоточной идентификации биологических генотипов. Список литературы:
1.
Уодингтон
К. Гомеорез // На пути к теоретической биологии. Т.1. Пролемены. М.:
Мир, 1970. С.21-39. 2.
Александров
В.Я. Клетки, макромолекулы и температура. Л.: Наука, 1978. 330 с. 62 3.
Инге-Вечтомов С. Т.
Неоднозначность матричных процессов как фактор адаптации II Системы надежности клетки,
Киев: Наук. думка, 1977. С.75-85. 4.
Александров В. Я.
Раативность клеток и белки. Л. : Наука, 1985. 318 с. 5. Richard Е., Сuellar - Glebnn A., Ford William F.T. Application of higher
derivative techniques to analysis of high resolution thermal denaturation
paoiiles of reassociated DNA // J.Boldii. 1978. Vol.75, N 13.
P.6026-6030. 6.
Королев А.М,
Исследований фотосинтетического
аппарата различных видов пшеницы и эгилопсов методом замедленной
флюоресценции //Сб.науч.тр. по прикладной ботанике, генетике и селекции
растений //ВИР. Л., I980. Т.67, вып.
2. с. 147-149. 7.
Гельфанд
И.М. , Гуфенкель В.С. и др. О
тактиках управления сложными системами в связи с физиологией. //
Биологические аспекты кибернетики. М.: Изд-во АН СССР, 1962, 330 с. 8.
Бирюков В. В. , К а я т е р в В.М.
Оптимизация периодических процессов микробиологического синтеза. М.:
Наука, 1985; 295 с. 9. Василенко C.М.
Применение математического планирования эксперимента для изучения
биохимических механизмов // Получение, исследование, применение антибиотиков
и биологически активных веществ: Тез. докл. международ. конф. молодых
ученых, 27-30 марта 1990 г. М., 1990.
с.21. 10. Той J.T., Gonsalez R.C. Patten
Recognition Principles. N.Y. 1974. 324 p. 11. Т
о u J.T., Computer and Information Sciences. III: Proc. of the Second
Symposim on Computer and Information Science. August 22-24, 1966. Academic,
H.Y. , 1967. p.34-39. 12. Ball G.H.,Hall J.P. IS0DATA.
An Iterative Method of Mulivariate Analysis and Pattern Classification.
Prooeedings of the IFJPS Congress, 1965. 13.
Скулачев
В.П. Энергетика биологических мембран. М.: Наука, 1989 564 с. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
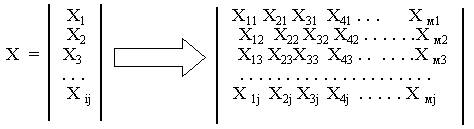
|
<<<
Interlibrary.narod.ru |
|
|